12nd Department of Obstetrics and Gynecology, University of Athens Medical School, Aretaieio Hospital, 21st Department of Obstetrics and Gynecology, University of Athens Medical School, Alexandra Hospital, Athens, 3Department of Obstetrics and Gynecology, University Hospital of Heraklion, Heraklion, Crete, 4Department of Obstetrics and Gynecology, University Hospital of Alexandroupolis, Democritus University of Thrace, Alexandroupoli, 5Department of Obstetrics and Gynecology, Hippokratio Hospital, Aristotle University of Thessaloniki, Thessaloniki, 6 Department of Obstetrics and Gynecology, Medical School, University of Thessalia, Larissa, Greece
Considering that preterm birth accounts for about 6-10% of all births in Western countries and of more than 65% of all perinatal deaths, elucidation of the particularly complicated mechanisms of labor is essential for determination of appropriate and effective therapeutic interventions. Labor in humans results from a complex interplay of fetal and maternal factors, which act upon the uterus to trigger pathways leading gradually to a coordinated cervical ripening and myometrial contractility. Although the exact mechanism of labor still remains uncertain, several components have been identified and described in detail. Based on the major role played by the human placenta in pregnancy and the cascade of labor processes activated via placental mediators exerting endocrine, paracrine, and autocrine actions, this review article has aimed at presenting the role of these mediators in term and preterm labor and the molecular pathways of their actions. Some of the aforementioned mediators are involved in myometrial activation and preparation and others in myometrial stimulation leading to delivery. In the early stages of pregnancy, myometrial molecules, like progesterone, nitric oxide, and relaxin, contribute to the retention of pregnancy. At late stages of gestation, fetal hypothalamus maturation signals act on the placenta causing the production of hormones, including CRH, in an endocrine manner; the signals then enhance paracrinically the production of more hormones, such as estrogens and neuropeptides, that contribute to cervical ripening and uterine contractility. These molecules act directly on the myometrium through specific receptors, while cytokines and multiple growth factors are also produced, additionally contributing to labor. In situations
leading to preterm labor, as in maternal stress and fetal infection, cytokines trigger placental signaling sooner, thus leading to preterm birth.
Activin, Corticotrophin releasing hormone (CRH), Follistatin, Inhibin, Interleukin-1 (IL-1), Neuropeptide Y (NPY), Neurokinine B (NKB), Oxytocin, Placental mediators, Preterm labor, Prostaglandins, Transforming growth factor α (TGF-α)
1.
INTRODUCTION
The human
placenta plays a major role in pregnancy, not only transporting gas and
nutrients from mother to fetus, but also producing brain, pituitary, gonadal
and adrenocortical peptides, and steroid hormones, chemically identical to and
biologically as active as their counterparts, along with cytokines and several
growth factors. Moreover, these molecules usually exert multiple functions
implicated in distant endocrine effects, cell to cell paracrine communication
or effects on the same cells (autocrine effects). Via these mediators, the
placenta is able to provide a favorable environment for the growing fetus
during pregnancy and to initiate the cascade of events that lead to labor.1
The aim of
this review article is to describe the role of placental mediators in term and
preterm labor and the molecular pathways of their actions. We examined review
articles and original research reports from 1990-2011 indexed in Pubmed and
other medical databases to present an updated overview of placental factors
involved in the activation of term and preterm labor and to elucidate their
biological plausibility and mode of action via a critical appraisal. The
delineation of these complicated mechanisms is the first step towards the
ultimate goal, this being the prevention of preterm labor, which still
complicates 10% of all pregnancies with short- and long-term consequences for
the fetus and neonate and also comprises a major cause of perinatal morbidity
and mortality.
The main placental mediators, their
actions, and cross actions are reviewed. We discuss the pathway of human labor
that is known to date, placing special emphasis on the additional or
supplementary pathways that, in the case of preterm labor, trigger these events
sooner than expected. Some actions are well established and others can be
hypothesized by using experimental data, such as receptor topography.
2. PLACENTAL MEDIATORS
Placenta products can be divided into five
categories: a) steroid hormones, b) hypothalamic-pituitary hormones, c)
neuropeptides, d) growth factors, e) cytokines.2,3 The actions of
mediators playing a main role in term and preterm labor are summarized in Table
1. Interestingly, some molecules are inducers for others possessing the same
actions, thus enhancing the cascade of labor. Moreover, most of the molecules
have multiple receptors in different sites of the maternal-placental-fetal unit
as well as exerting complex endocrine, paracrine, and autocrine effects on
those sites.
2.1 Placental steroid hormones
Steroid hormones produced by the placenta
include the common female gonadal steroid hormones, progesterone and estrogens,
mainly estriol, but also estradiol and estrone. The predomination of estrogen
over progesterone occurs during the transition from early pregnancy to labor
and delivery in humans. In other mammals the production of progesterone is
decreased and is partially converted to estrogen via the enzyme 17-hydroxylase.
The human placenta lacks this enzyme, thus fetal dehydroepiandrosterone sulfate
(DHEA-S) comes into play for the production of estrogen.1 The
converting placental enzymes are 3-hydroxysteroid dehydrogenase and delta 4-5
isomerase. With progesterone receptors achieving desensitization and increased
estrogen levels at term, estrogen-dependent molecules, like receptors and
enzymes, induce labor.
2.2 Hypothalamic-pituitary hormones
produced by the placenta
It is
noteworthy that the placenta produces hormones that are known to be produced by
neuroendocrine organs, such as the hypothalamus and pituitary gland. These
hormones include human chorionic gonadotrophin (hCG), human placental lactogen
(hPL), oxytocin, gonadotrophin-releasing hormone (GnRH), opioids, growth
hormone-releasing hormone (GHRH), somatostatin, and corticotrophin releasing
hormone (CRH).
GnRH stimulates hCG placental production
which is induced by estrogen, cytokines, and other molecules, whereas hCG
production is blocked by progesterone, opioids, and other hormones. This may
explain why recurrent miscarriages occur in opioid-addicted pregnant women.2
Oxytocin placental production and oxytocin
receptors are increased during pregnancy. Oxytocin acts on myometrium and
decidual tissues resulting in uterine contractions and prostaglandins
production, respectively.4
Corticotropin-releasing factor-like
activity in human placental extracts has been reported since the 1980's.5,6
Women who go into preterm labor have higher levels of plasma CRH as early as
15-16 weeks compared with women who deliver at term.2,7 On the other
hand, women delivering post-term have lower levels of plasma CRH.1
Lower CRH production early in pregnancy has utero-relaxing action, but these
early CRH receptors later in pregnancy and near labor are desensitized by a
kinase which is oxytocin-controlled.8 Before labor other "late" CRH
receptors are expressed in fetal pituitary, fetal membranes, the deciduas, and
the myometrium, which activate G-protein mediated pathways that lead to
prostaglandins production and myometrial contractions.9,10
Furthermore, the CRH-binding protein (CRH-BP), a 37kD protein produced in the
placenta, liver, brain, and other sites, blocks the action of CRH early in
pregnancy. The binding of CRH to CRH-BP makes it biologically inactive and
removes it from the circulation. Near term or in preterm labor, CRH-BP
decreases coincidentally with the increase of CRH production, this enhancing
its action.11,12
Very recently, molecules belonging to the
family of CRH, named urocortins, have been described. They bind to the same
receptors as CRH, their levels increase before labor, and they result in
myometrial contractility in the same way, directly and indirectly, as CRH.11,13
2.3 Placental neuropeptides
Two known neuropeptides produced by the
placenta are neuropeptide Y (NPY) and neurokinine B (NKB). In humans, during
pregnancy the placenta secretes NPY in the maternal and fetal circulation and
in the amniotic fluid. Throughout gestation NPY levels do not change
significantly; however, during labor NPY levels in all the above biological
fluids progressively increase, with the highest levels occurring at the most
advanced stages of labor, the latter verification confirming the role of NPY in
labor and delivery. Immediately after delivery, NPY levels fall, this
demonstrating its placental origin. NPY receptors are present in all peripheral
cells of placental terminal villi, thus indicating that NPY possibly acts both
ways: that is, there may be an enhancement of CRH production in an autocrine
manner. Furthermore, NPY receptors also are identified in human myometrium. NPY
binding on myometrial receptors contributes to uterine contractility through a
pertussis toxin sensitive G-protein mediated pathway. This pathway starts from
the NPY receptors and continues with inhibition of adenylate cyclase activity
and increase of intracellular calcium levels (Figure 1). In vitro studies have
shown involvement of NPY in regulation of uterine blood flow and uterine
contractility.11
NKB, another neuropeptide produced by the
placenta, acts as a paracrine regulator of fetal placental vascular tone mainly
causing relaxation of placental vessels. Moreover, it is a modulator of
myometrial contractility, enhancing it when binding with its receptors (NKR) on
myometrium. NKB concentration in maternal blood increases as pregnancy advances
and decreases rapidly after delivery, this demonstrating its placental origin.
NKB receptors belong to the G-protein-linked receptors family. NKB initiated
pathways are similar to NPY receptor initiated pathways (Figure 2). Expression
of placental NKB mRNA and of protein is higher in term labor than at elective
cesarean section and even higher in preterm labor.11,14
2.4 Placental growth factors
The placenta is the source of numerous
growth factors including inhibin, activin, follistatin, transforming growth
factor α (TGF-α), insulin-like growth factor (IGF), epidermal growth factor
(EGF), fibroblast growth factor (FGF), platelet-derived growth factor (PDGF),
and angiogenin.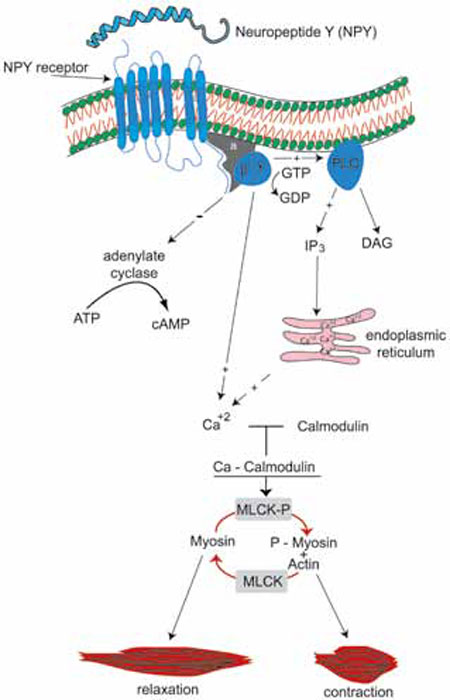
Figure 1. NPY binding to its receptors contributes to uterine contractility through a pertussis toxin sensitive G-protein mediated pathway. This pathway starts from the NPY receptors and continues with inhibition of adenylate cyclase activity and increase of intracellular calcium levels. 
Figure 2. NKB receptors belong to the G-protein-linked receptors family. The NKR initiated pathway is similar to the NPY receptor pathway. The result of its action is an increase of intracellular calcium levels that enhance myometrial contractility. The difference of its action is the induction and not inhibition of a denylate cyclase.
Activin A is a disulfide-linked dimer that
belongs to the transforming growth factor β (TGF-β) superfamily. An increase of
placental activin A production occurs in labor at term. Increased production
also occurs at 31-34 weeks of gestation in cases that will go into preterm
labor, but there is no increase during preterm labor, although there is
increased activin A mRNA expression at that time.15,16 Activin A
enhances the production of prostaglandins and oxytocin in a paracrine manner in
the placenta. Administration of inflammatory cytokines to gestational tissue
was reported to result in increased activin A production, which may be relevant
in cases of preterm delivery occurring secondarily to intrauterine infection.17
Activin A receptors are transmembrane heteromeric serine kinases that interact
with and phosphorylate cytoplasmic SMAD2, which multimerizes with SMAD4 and
translocates to the nucleus where these proteins can act as transcription
factor complexes (Figure 3).11 The SMAD proteins are homologs of
both the drosophila protein, mothers against decapentaplegic (MAD), and the
Caenorhabditis elegans protein SMA, the name being a combination of the two
aforementioned proteins. SMADs are intracellular proteins that transduce
extracellular signals from TGF-β ligands to the nucleus where they activate
downstream TGF-β gene transcription.
Figure 3. Activin A receptors are transmembrane heteromeric serine kinases that interact with and phosphorylate cytoplasmic SMAD2, which multimerizes with SMAD4 and translocates to the nucleus, where these proteins can act as transcription factor complexes.
Inhibin and follistatin both inhibit
activin A, but their constant presence from early on and during pregnancy
minimizes the possibility of their playing a role in labor (Figure 4),2,15
Another growth factor, angiogenin, displays increased second trimester amniotic
fluid levels in pregnancies with higher risk of spontaneous preterm delivery.18
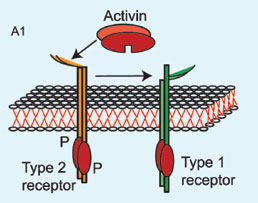
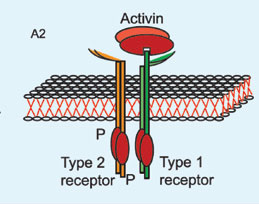

Figure 4. The inhibitory action of follistatin on activin. A1 and A2: Activin interaction with type 2 receptor enhances binding of type 1 receptor and formation of the active heteromeric receptor. B. Follistatin binds with activin so that the complex molecule cannot interact with activin type 2 receptor and the active heteromeric receptor cannot be formatted.
Other less studied placental growth
factors are EGF, IGF, and TGF-α. IGF production increases throughout pregnancy:
though studies suggest that IGF mediates stretch-induced myometrial
hypertrophy, further investigation is needed to precisely determine their
possible role in labor.19 EGF and TGF-α seem to share the same
receptors on the placenta, endometrium, and decidual and fetal membranes.
Studies indicate that there is upregulation of epidermal growth factor
receptors messenger RNA expression in myometrium and fetal membrane before and
after the onset of labor.20 EGF and TGF-α also induce prostaglandin
synthesis, but there is no significant increase in their production before
labor, thus their role in labor is still in question.2,10
2.5 Cytokines produced by placental
leukocytes
White blood cells located in the placenta
secrete cytokines, including interleukin 1 (IL-1), interleukin 6 (IL-6), tumor
necrosis factor α (TNF-α), and interferon (IFN). Inflammatory cytokines, like
IL-1 and TNF-α, induce prostaglandins production and may thus account for
preterm delivery in the setting of infection.10,21,22
Bacterial toxins, e.g. LPS
(lipopolysaccharide S), act on decidual tissue to cause the production of TNF-α
which acts in the amnion or the decidua to alter cell function, namely, cell
growth or prostaglandin production or both. A variety of proteins produced in
the decidua accumulate in the amniotic fluid. TNF-α, however, is not present in
amniotic fluid of normal pregnancies at any stage of gestation or during spontaneous
labor at term. However, TNF-α is present in amniotic fluid of 4 out of 10
pregnancies complicated by preterm labor in which the membranes are intact; LPS
is also present in the amniotic fluid. We cannot be sure that TNF-α is not
produced in the decidua in some preterm labor pregnancies in which LPS is
present in the amniotic fluid, although TNF-α has not to date been identified.
In 6 out of 10 amniotic fluids in which LPS was present, TNF-α was not
detected.23 This could mean that TNF-α was not produced or else too
little was transferred and accumulated in amniotic fluid to be detected. It is
also observed that TNF-α is cytostatic, but not cytolytic, in human amnion
cells. Amnion cells, therefore, are relatively unique because other normal
cells commonly respond to TNF-α as a growth factor or, alternatively, they do
not. Indeed, TNF-α, while cytolytic or cytostatic in a number of tumor cells,
usually does not inhibit growth in normal cells. The possibility thus exists
that in the amnion, LPS or some other bacterial factor acts on macrophage-like
decidual cells (or macrophages of the decidua or both) to cause the production
of TNF-α, which, in turn, may act to inhibit growth of the amnion, a process
that may favor rupture of the membranes. TNF-α may also act in vivo, as it does
in vitro, to stimulate the production of PGF2α by the decidua and
PGE2 by the amnion.
The decidua,
as well as endometrial stromal cells, also produces IL-1. The concentration of
IL-1 in amniotic fluid increases during spontaneous parturition and increases
even more dramatically during preterm labor complicated by infection. IL-1 acts
on the decidua and endometrial stromal cells to augment PGF2α
formation.23 Moreover, IL-1 and TNF-α stimulate the activity of the
transcriptional factor NF-κB (Nuclear Factor κB) in the uterus. NF-κB plays a
significant role in "functional" progesterone withdrawal, which is required for
the initiation of labor. NF-κB also stimulates prostaglandin synthesis and
regulates matrix metalloproteinase expression, connexin-43, and oxytocin
receptor. IL-1 additionally induces production of CRH receptors and seems to
have a synergic action with EGF.10
Other
cytokines with a possible role in preterm labor are granulocyte colony
stimulating factor, IL-6, and IL-8.3 IL-1 and IL-6 enhance
endometrial CRH gene expression.24 IL-1 induces CRH receptor
expression and function, contributing to a preparatory contractile environment
necessary for the initiation of labor. IL-6 also induces production of
metalloproteinases that promote cervical ripening and rupture of membranes, as
observed in ex vivo studies.25
3. LABOR AND DELIVERY
Throughout
most of pregnancy, some low frequency, weak, and poorly coordinated
contractions (Braxton-Hicks) occur. Coupling of actin and myosin prompts the
occurrence of these contractions, this coupling resulting from phosphorylation
of myosin by the enzyme myosin light-chain kinase (MLCK). The activation of
MLCK is achieved via calmodulin, which is activated by intracellular calcium.
Progesterone, prostacyclin, nitric oxide, relaxine, parathyroid hormone-related
peptide, and other mediators increase cyclic adenosine or guanosine
monophosphate molecules (cAMP or cGMP), which in turn inhibit the release of
calcium from intracellular deposits.3
Moreover,
there are no connexin gap junctions between myometrial cells during the period
of Braxton Hicks contractions.26 Connexins, and mainly connexin 43
expression, are uterine stretch-induced and estrogen-induced,27
while progesterone blocks this expression. With progesterone receptors
desensitization and increased estrogen levels at term of pregnancy, connexin
gap junctions are massively increased.
The role of the placenta, by means of
endocrine and paracrine mediators, is the synchronization of fetal maturation
with parturition.10 Parturition has two stages, myometrial
activation (stage 1) and myometrial stimulation (stage 2). Myometrial
activation is achieved via myometrial oxytocin and prostaglandins receptors
synthesis and induction of myometrial cells gap junctions in response to
estrogen.10 Myometrial stimulation comes about through oxytocin and
prostaglandins at the time that they bind to their receptors, with or without
the combined action of mediators like neuropeptides and cytokines.
3.1 Progesterone withdrawal in labor
Ιn humans, progesterone levels remain high
throughout pregnancy and during labor. This has led to the hypothesis of a
"functional" progesterone withdrawal, mentioned above. Τhere are two major
isoforms of progesterone receptor, PR-A and PR-B, which belong to the nuclear
receptor superfamily, as well as many other isoforms which have so far given
evidence of being less significant. PR-B is the main receptor for the nuclear
signal transduction of progesterone. The expression of less sensitive isoforms
possibly contributes to the "functional" withdrawal of progesterone before
labor.28,29
The binding of progesterone to PR-C, which
is a soluble form of the receptor, may sequester available progesterone away
from PR-B and thereby diminish its biological effect. In a study of women in
labor, PR-C protein levels were increased in cytoplasmic fractions of fundal
myometrial cells. A reduction of progesterone nuclear coactivators has also
been noticed pointing to the "closing" of chromatin structure and the reduced
expression of PR-responsive genes as a possible mechanism for the initiation of
labor.28 Finally, transcriptional factor NF-κB, which is involved in
the synthesis of a large number of proinflammatory mediators such as cytokines,
antagonizes progesterone in reproductive tissues during labor.28
3.2 Placental CRH and the fetal
hypothalamic-pituitary-adrenal axis
The increase of oxytocin and
prostaglandins receptors is signaled by the fetal
hypothalamic-pituitary-adrenal (HPA) axis.30 Once fetal maturity is
reached, the fetal hypothalamus and placenta secrete CRH, which releases
adrenocorticotropic hormone (ACTH) from fetal pituitary and fetal adrenals
which produce cortisol and DHEA-S, the latter being aromatized to estrogens by
the placenta.12 Fetal cortisol is essential for maturation of fetal
organs, and especially the respiratory system, needed for extrauterine life.
Increased cortisol levels promote the production of prostaglandins from the
placenta, through prostaglandin synthetase (PGS), in an estrogen-dependent
manner.8,22,31 In sheep, placenta prostaglandins stimulate P450,
resulting in increased placental estrogen synthesis.8
Placental CRH synthesis is stimulated by
the produced fetal cortisol (positive feedback mechanism).10 A
difference exists between placental and hypothalamic CRH, since placental
secretion is stimulated and fetal hypothalamic secretion is depressed by
cortisol. Thus, although fetal CRH production may be reduced, placental CRH is
increased. Moreover, the potential for fetal cortisol negative feedback action
on fetal ACTH production is reduced by increased production of
corticosteroid-binding globulin (CBG) at the end of pregnancy. Placental
estrogens also induce the expression of the PGS and, in turn, the production of
prostaglandins in chorion and amnion cells. Furthermore, placental estrogens
enhance placental CRH production, this forming a second positive feedback loop.10
Prostaglandin dehydrogenase (PGDH) is
induced by progesterone and downregulated by cortisol. 31
Progesterone "functional" withdrawal reduces the activity of progesterone and
increases the level of expression of estrogen receptors and estrogen activity
on the myometrium. As a result, there is an increase in production of
estrogen-dependent molecules, such as oxytocin receptors and prostaglandins,
which promote uterine contractility. Activated prostaglandins receptors elevate
intracellular calcium by blocking cAMP. They also enhance the production of
cervix metalloproteinases to promote cervical ripening and decidual and fetal
membrane activation.8 The initiation of this cascade seems to be
placental production of CRH.32
3.3 Prostaglandins in labor and delivery
There is considerable evidence in support
of the likelihood that prostaglandins serve as the mediators (uterotonins) of
the myometrial contractions experienced by women in labor. This evidence can be
summarized as follows: (a) the administration of prostaglandins
(intraamniotically, vaginally or intravenously) in human pregnancy leads to the
onset of myometrial contractions and delivery at any stage of gestation; (b)
the administration of inhibitors of prostaglandin synthesis to pregnant women
is associated with prolonged gestation and an increase in the time interval for
intraamniotic hypertonic saline induction of abortion; and (c) the
concentrations of prostaglandins in amniotic fluid and in maternal plasma and
urine are increased during parturition. The levels of PGE2 and PGF2α
in amniotic fluid increase strikingly during labor.23
Prostaglandins, mainly PGE2 and to a
lesser extent PGF2α, seem to be the endpoint of the CRH cascade and
characterize the action of most of the molecules that participate in term and
preterm labor mechanisms. However, they also enhance their own production. They
decrease the activity of placental 11β-HSD-2, the enzyme that converts
cortisol, resulting in a further increase in local cortisol concentrations,
this representing another positive feedback mechanism.8 Moreover,
PGE2 stimulates fetal CRH production, yet a further positive feedback loop.31
A remarkable feature of prostaglandins is
that when they bind with receptors type 1 and 3, which exist in the upper
segment of the uterus, they cause contractility, but when they bind with
receptors types 2 and 4, which exist in the lower segment of the uterus, they
cause relaxation, a heterogeneity of the receptors that is genetically
controlled. This constitutes a well established part of the mechanism of labor.
The mechanisms discussed above are shown in Figure 5.
4. PRETERM LABOR
The greatest single cause of newborn
morbidity and mortality is preterm birth.33,34 Well known common
causes leading to preterm birth include stress, systemic or maternal genital
tract infections, placental ischemia or vascular lesions, and uterine
overdistension.35,36 It is reasonable to hypothesize that under
these circumstances the placental "clock" is triggered sooner than normal to
initiate the cascade of events leading to labor.
4.1 Stress
The placenta is responsive to maternal
stress.37 CRH is released from cultured human placental cells in a
dose-dependent manner in response to all major biological effectors of stress,
such as cortisol, catecholamines, oxytocin, angiotensin 2, and IL-1.7,10
Thus, in preterm delivery it may be the maternal HPA axis and the sympatho-adrenergic
system that starts the placental CRH cascade. Activation of the fetal HPA axis
can occur in an adverse intrauterine environment, like compromised
uteroplacental blood flow and other conditions of fetal hypoxemia or infection.8,38
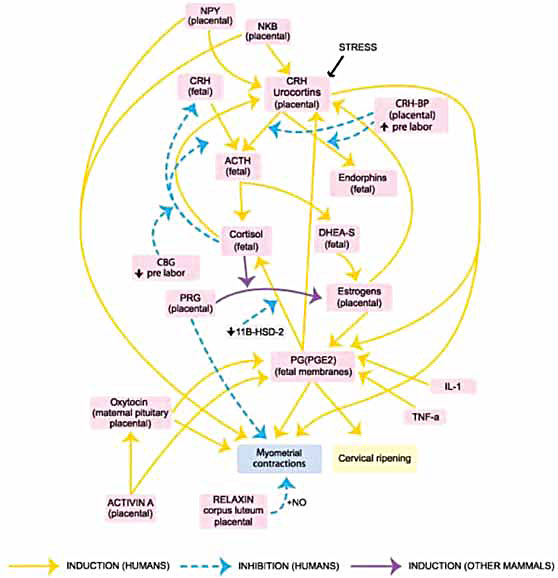
Figure 5. Pathways leading to labor. The main pathway starts from placental CRH, exploits fetal DHEA-S to produce estrogen environment and, through prostaglandins, results in myometrial contractions and cervical ripening, thus inducing labor. A variety of mediators interact in complex ways producing positive feedback loops of the main pathway:
- placental CRH → fetal ACTH → fetal cortisol → placental CRH
- placental CRH → fetal ACTH → fetal DHEA-S → estrogens → placental CRH
- placental CRH → fetal ACTH → fetal DHEA-S → estrogens → PGE2 → placental CRH
- placental CRH → fetal ACTH → fetal DHEA-S → estrogens → PGE2 → fetal cortisol → placental CRH.
These mediators feed-forward enhancing roots of the main pathway (NPY, NKB, IL-1, TNF-α, stress mediators, like catecholamines) or the endpoint of labor (NPY, NKB, oxytocin, activin A, IL-1, TNF-α).
4.2 Infection and inflammation
Microarray and suppression-subtractive
hybridization studies have demonstrated an upregulation of a vast number of
proinflammatory genes in fetal membranes, myometrium, and the cervix with the
onset of labor.39 Such upregulation is demonstrated by an influx of
inflammatory cells into the uterus and increased expression of proinflammatory
cytokines.40
At term, this inflammatory response may be
triggered by the maturing fetal lung, which produces increased surfactant lipid
and protein, leading to activation of macrophages in the amniotic fluid and
increased inflammatory response in the uterus.41
Bacterial toxins like LPS, originating in
microorganisms in uterine or extrauterine tissues, may act on the
macrophage-like decidua to initiate a series of events that culminate in
preterm labor, possibly together with premature rupture of the fetal membranes.
Evidence in support of this hypothesis is that more than 40 years ago it was
shown that the administration of LPS to pregnant animals caused abortion or
premature parturition. Moreover, in LPS-treated animals, abortion or preterm
delivery is associated with decidual hemorrhage and necrosis. Finally, LPS acts
on monocytes and macrophages to instigate the production of prostaglandins,
TNF-α, and IL-1.23
Inflammation is a critical trigger for the
initiation of term and preterm labor, with NF-κΒ being a transcriptional factor
implicated in proinflammatory stimuli signaling. IL-1 and TNF-α stimulate NF-κB
activity in the uterus. NF-κB also stimulates prostaglandin synthesis and
regulates matrix metalloproteinase expression, connexin-43, and the oxytocin
receptor. As already mentioned, NF-κB is also related to a "functional"
progesterone withdrawal in the human myometrium.28
Inflammatory cytokines, like IL-1 and
TNF-α, also induce prostaglandin production via upregulation of PGS synthetase
and downregulation of PGDH and thus may explain preterm delivery in the setting
of infection.10,21,22
An interesting study demonstrated
synthesization and secretion of TNF-α into the culture medium by human decidual
cells and explants in response to treatment with LPS. LPS treatment also caused
an increase in PGF2α production by decidual cells and explants.23
Other cytokines with a possible role in
preterm labor are granulocyte colony stimulating factor, IL-6, and IL-8.
[5] Meanwhile, CRH receptors have been identified in the
syncytiotrophoblast, deciduas and endometrium. PGE2, IL-1, and IL-6 enhance
endometrial CRH gene expression and IL-1 induces CRH receptor expression and
function, contributing to a preparatory contractile environment essential for
the initiation of labor. Nevertheless, when an infection is present the fetal
HPA axis can also be activated, thus starting the cascade.42,43
Recently, studies have shown a significant
impact of chorioamnionitis on placental CRH and urocortins and their receptors,
suggesting that placental expression of stress-related pathways is activated in
infective processes.44 Even cervical incompetence manifesting as a
case of preterm labor, mechanical causes excluded, may be related to infection
and inflammation, as a shortened or damaged cervix would not be capable of
impeding the ascent of bacteria.
4.3 Abruption and ischemia
The key molecule in preterm labor when
placental abruption and ischemia is present, as in placental thrombosis, is
thrombin. It has been determined that thrombin binds to receptors which
increase intracellular calcium via G-protein coupling and phospholipase C
activation.45 In vitro, and moreover in vivo, experiments have shown
a significant reduction of myometrial contractility via administration of the
thrombin inhibitor heparin.46 Finally, thrombin induces the
production of metalloproteinases, which can cause cervical ripening and rupture
of fetal membranes, and protease-activated receptors which stimulates
myometrial contractions.47,48
4.4 Overdistention
In cases of uterine stretch, like multiple
gestation or polyhydramnios, myometrial and amnion epithelial cells increase
their expression of cyclooxygenase and prostaglandin synthesis. Again, NF-κB
seems to be the key signaling element.49
5. CONCLUSION
Human placenta is seen to be both an
intermediary barrier and an active messenger in the maternal-fetal dialog. It
releases endocrine, paracrine, and autocrine factors which control secretion of
regulatory or terminal-effector molecules. Coordinated action results in
cervical ripening, myometrial contractility, and labor, placenta also being
implicated in initiation of labor and in preterm labor. Meanwhile, placental CRH
acts on the fetal pituitary-adrenal axis, stimulating adrenal production of
DHEA-S and cortisol, while myometrial activation, achieved via activation of
estrogen derived from fetal DHEA-S through placental enzymes, causes uterine
contractions. Subsequently, with onset of late-gestation "functional"
progesterone withdrawal, fetal cortisol production, developing in an estrogenic
environment, leads to labor. Placental CRH also acts directly on myometrial
cells via its receptors. Other placental mediators (e.g. neuropeptides produced
before labor), binding to their own receptors in myometrium, enhance uterine
contractility. Placental CRH-induced prostaglandin and molecules (e.g.
oxytocin, placental neuropeptides), regulatory cytokines, and growth factors cause
cervical ripening, rupture of fetal membranes, myometrial stimulation, and
regular contractions. Cytokines (e.g. IL-1, IL-6, and TNF-α) are involved in
positive feedback of placental CRH production, while stress or infection can
trigger the placental "clock" and prematurely initiate the cascade of labor.
Despite extensive research into the
extremely complicated mechanisms of labor in numerous animal models and humans,
full elucidation is still lacking because of broad human v. animal divergence
of physiology. Future progress in this area will enable development of
effective treatments for preterm delivery thereby reducing neonatal morbidity
and mortality.
REFERENCES
1. Federenko IS, Wadhwa PD, 2004 Women's Mental Health During Pregnancy Influences Fetal and Infant Developmental and Health Outcomes. CNS Spectrums 9: 198-206.
2. Petraglia F, Florio P, Nappi C, Genazzani AR, 1996 Peptide Signaling in Human Placenta and Membranes: Autocrine, Paracrine and Endocrine Mechanisms. Endocr Rev 17: 156-186.
3. Reis FM, D'Antona D, Petraglia F, 2002 Predictive value of hormone measurements in maternal and fetal complications of pregnancy. Endocr Rev 23: 230-257.
4. Fuchs AR, Fuchs F, Husslein P, Soloff MS, Fernstrom MJ, 1982 Oxytocin receptors and human parturition: a dual role for oxytocin in the initiation of labor. Science 215: 1396-1398.
5. Shibasaki T, Odagiri E, Shizume K, Ling N, 1982 Corticotropin-releasing factor-like activity in human placental extracts. J Clin Endocrinol Metab 55: 384-386.
6. Frim DM, Emanuel RL, Robinson BG, et al, 1988 Characterization and gestational regulation of corticotrophin-releasing hormone messenger RNA in human placenta. J Clin Invest 82: 287-292.
7. McLean M, Smith R, 1999 Corticotropin-releasing hormone in human pregnancy and parturition. Trends Endocrinol Metab 10: 174-178.
8. Challis JRG, Sloboda DM, Alfraidy N, et al, 2002 Prostaglandins and mechanisms of preterm birth. Reproduction 124: 1-17.
9. Karteris E, Grammatopoulos D, Randeva H, Hillhouse EW, 2000 Signal transduction characteristics of the corticotropin-releasing hormone receptors in the feto-placental unit. J Clin Endocrinol Metab 85: 1989-1996.
10. Carbillon L, Uzan M, Challier JC, Marviel P, Uzan S, 2000 Fetal-placental and decidual-placental units: role of endocrine and paracrine regulations in parturition. Fetal Diagn Ther 15: 308-318.
11. Torricelli M, Giovanelli A, Torres PB, et al, 2006 Endocrine and paracrine mechanisms in human placenta and fetal membranes in preterm birth. Preterm Birth 3: 35-41.
12. Reis FM, Fadalti M, Florio P, Petraglia F, 1999 Putative role of placental corticotrophin-releasing factor in the mechanisms of human parturition. J Soc Gynecol Investig 6: 109-119.
13. Pepels PP, Spaanderman ME, Bulten J, et al, 2009 Placental urocortins and CRF in late gestation. Placenta 30: 483-490.
14. Torricelli M, Giovannelli A, Leucci E, et al, 2007 Placental neurokinin B mRNA expression increases at preterm labor. Placenta 28: 1020-1023.
15. Keelan JA, Marvin KW, Sato TA, et al, 1999 Concentrations of activin A, inhibin A and follistatin in human amnion, choriodecidual and placental tissues at term and preterm. J Endocrinol 163: 99-106.
16. Plevyak MP, Lambert-Messerlian GM, Farina A, Groome NP, Canick JA, Silver HM, 2003 Concentrations of serum total activin A and inhibin A in preterm and term labor patients: a cross-sectional study. J Soc Gynecol Investig 10: 231-236.
17. Keelan JA, Zhou RL, Evans LW, Groome NP, Mitchell MD, 2000 Regulation of activin A, inhibin A, and follistatin production in human amnion and choriodecidual explants by inflammatory mediators. J Soc Gynecol Investig 7: 291-296.
18. Farina A, Carinci P, Di Luzio L, 1997 Angiogenin: a marker for preterm delivery in midtrimester amniotic fluid. Am J Obstet Gynecol 177: 483-484.
19. Shynlova O, Tsui P, Dorogin A, Lowell-Langille B, Lye SJ, 2007 Insulin-like growth factors and their binding proteins define specific phases of myometrial differentiation during pregnancy in the rat. Biol Reprod 76: 571-578.
20. Chen J, Zenq W, Wu J, 2000 Correlation between epidermal growth factor receptor messenger RNA expression and the onset of labor. Zhonghua Fu Chan Ke Za Zhi 35: 29-31.
21. Bowen JM, Chamley L, Keelan JA, Mitchell MD, 2002 Cytokines of the placenta and extra-placental membranes: roles and regulation during human pregnancy and parturition. Placenta 23: 257-273.
22. Challis JRG, Lye SJ, Gibb W, Whittle W, Patel F, Alfaidy N, 2006 Understanding preterm labor. Ann NY Acad Sci 943: 225-234.
23. Casey ML, Cox SM, Beutler B, Milewich L, MacDonald PC, 1989 Cachectin/Tumor Necrosis factor-a formation in human decidua - potential role of cytokines in Infection-induced preterm labor. J Clin Invest 83: 430-436.
24. Vitoratos N, Mastorakos G, Kountouris A, Papadias K, Creatsas G, 2007 Positive association of serum interleukin-1beta and CRH levels in women with pre-term labor. J Endocrinol Invest 30: 35-40.
25. Lee SY, Buhimschi IA, Dulay AT, et al, 2011 IL-6 trans-signaling system in intra-amniotic inflammation, preterm birth, and preterm premature rupture of the membranes. J Immunol 186: 3226-3236.
26. Garfield RE 1988 Structural and functional studies of the control of myometrial contractility and labour. In McNellis D, Challis JRG, MacDonald PC, Nathanielsz PW, Roberts JM (eds) The onset of labour: cellular and integrative mechanisms. Ithaca, NY: Perinatology Press; pp, 55-80.
27. Lye SJ, Ou CW, Teoh TG, et al, 1998 The molecular basis of labour and tocolysis. Fetal Matern Med Rev: 121-136.
28. Vrachnis N, Malamas FM, Sifakis S, Tsikouras P, Iliodromiti Z, 2012 Immune aspects and myometrial actions of progesterone and CRH in labor. Clin Dev Immunol 2012: 937618.
29. Vrachnis N, Malamas FM, Sifakis S, Deligeoroglou E, Iliodromiti Z, 2011 The oxytocin-oxytocin receptor system and its antagonists as tocolytic agents. Intl J Endocrinol 2011: 350546.
30. Liggins GC, Thorburn GD, 1994 Initiation of parturition. In Lamming GE (ed) Marshall's: Physiology of Reproduction. London, UK: Chapman and Hall; pp, 863-1002.
31. Challis JRG, Smith SK, 2001 Fetal endocrine signals and preterm labor. Biol Neonate 79: 163-167.
32. Gibb W, Challis JR, 2002 Mechanisms of term and preterm birth. J Obstet Gynaecol 24: 873-883.
33. Rush RW, Keirse MJNC, Howat P, Baum JD, Anderson AB, Turnbull AC, 1976 Contribution of preterm delivery to perinatal mortality. Br Med J 2: 965-968.
34. Vrachnis N, Karavolos S, Iliodromiti Z, et al, 2012 Impact of mediators present in amniotic fluid on preterm labour. In Vivo 26: 799-812.
35. Vrachnis N, Vitoratos N, Iliodromiti Z, Sifakis S, Deligeoroglou E, Creatsas G, 2010 Intrauterine inflammation and preterm delivery. Ann NY Acad Sci 1205: 118-122.
36. Malamitsi-Puchner A, Vrachnis N, Samoli E, et al, 2006 Investigation of midtrimester amniotic fluid factors as potential predictors of term and preterm deliveries. Mediators Inflamm 2006: 94381.
37. Wadhwa PD, Culhane JF, Rauh V, Barve SS, 2001 Stress and preterm birth: Neuroendocrine, immune/inflammatory, and vascular mechanisms. Matern Child Health J 5: 119-125.
38. Vrachnis N, Malamitsi-Puchner A, Samoli E, et al, 2006 Elevated mid-trimester amniotic fluid ADAM-8 concentrations as a potential risk factor for preterm delivery. J Soc Gynecol Investig 13: 186-190.
39. Charpigny G, Leroy MJ, Breuiller-Fouche M, et al, 2003 A functional genomic study to identify differential gene expression in the preterm and term human myometrium. Biol Reprod 68: 2289-2296.
40. Young A, Thompson AJ, Ledingham M, et al, 2002 Immunolocalisation of proinflammatory cytokines in myometrium, cervix and fetal membranes during human parturition at term. Biol Reprod 66: 445-449.
41. Condon JC, Jeyasuria P, Faust JM, et al, 2004 Surfactant protein secreted by the maturing mouse fetal lung acts as a hormone that signals the initiation of parturition. Proc Natl Acad Sci USA 101: 4978-4983.
42. Gravett MG, Haluska GJ, Cook MJ, Novy MJ, 1996 Fetal and maternal endocrine responses to experimental intrauterine infection in rhesus monkeys. Am J Obst Gynecol 174: 1725-1733.
43. Gravett MG, Hitti J, Hess DL, Eschenbach DA, 2000 Intrauterine infection and preterm delivery: Evidence for activation of the fetal hypothalamic-pituitary-adrenal axis. Am J Obst Gynecol 182: 1404-1413.
44. Torricelli M, Novembri R, Bloise E, De Bonis M, Challis JR, Petraglia F, 2011 Changes in placental CRH, urocortins, and CRH-receptor mRNA expression associated with preterm delivery and chorioamnionitis. J Clinl Endocrinol Metab 96: 534-540.
45. Bohm SK, McConalogue K, Kong W, Bunnett NW, 1998 Proteinase-activated receptors: new functions for old enzymes. News Physiol Sci 13: 231-240.
46. Elovitz MA, Ascher-Landsberg J, Saunders T, Phillippe M, 2000 The mechanisms underlying the stimulatory effects of thrombin on myometrial smooth muscle. Am J Obst Gynecol 183: 674-681.
47. Mackenzie AP, Schatz F, Krikun G, Funai EF, Kadner S, Lockwood CJ, 2004 Mechanisms of abruption-induced premature rupture of the fetal membranes: Thrombin enhanced decidual matrix metalloproteinase-3 (stromelysin-1) expression. Am J Obst Gynecol 191: 1996-2001.
48. Stephenson C, Lockwood C, Ma Y, Guller S, 2005 Thrombin-dependent regulation of matrix metalloproteinase (MMP-9) levels in human fetal membranes. Journal of Maternal-Fetal and Neonatal Medicine 18: 17-22.
49. Mohan AR, Soorana SR, Lindstrom TM, et al, 2007 The effect of mechanical stretch on cyclooxygenase type 2 and activator protein-1 and nuclear factor-kappa B activity in human amnion cells. Endocrinology 148: 1850-1857.
Address for correspondence:
Nikolaos Vrachnis, MD, 124B Vasilisis Sofias Av., 11526, Athens, Greece,
E-mail: nvrachnis@med.uoa.gr
Received 08-04-12, Revised 20-06-12, Accepted 10-08-12